

A Beginner’s Guide to Theropods, Part 5: Plant-eating Meat-eating Dinosaurs
Early in the Cretaceous, a revolution was afoot. Not in the continents or climate but in the organisms themselves. To modern eyes, Jurassic floras would have looked like an odd mix of familiar ferns and conifers with tropical relicts like cycads and ginkgos. Most conspicuous would be the absence of any flowering plants: there were no broadleaf trees, no fruits or nuts, and no social insects to pollinate them (Benton). As these new plants spread with the continents (Gurung et al), there came an array of new herbivores, each specially adapted for feeding on them: first iguanodonts and ankylosaurs, later ceratopsians and hadrosaurs. And radiating out alongside them was an unprecedented diversity of theropods, including, for the first time, a wide range of non-predators (Zanno and Mackovicky). Where pencil-toothed diplodocids and narrow-mouthed stegosaurs had previously scraped pine needles and cropped cycads, there were now therizinosaurs munching leaves, oviraptorosaurs crushing seeds, and alvarezsaurs digging up ants.
How smart were theropods?
How smart is any animal? It's impossible to really know, but we can make estimates. A popular one, the Encephalization Quotient, is obtained by plotting the brain-to-body mass ratio of one animal against a curve made by a range of differently-sized, similar animals. This is because, as animals get bigger, brains don't grow at the same rate but actually decrease relative to body size. As the number of neurons grows, connections increase exponentially, meaning fewer are required to perform the same basic functions. For instance, an elephant has a brain-to-body mass ratio about 14 times smaller than that of a mouse! One study (Hubert et al), comparing them to various birds and reptiles, estimated ornithomimids and maniraptorans to be “brainier” than all other theropods, within the low-end range of living birds and well above average for crocodilians.
Of course, size alone isn't the only factor determining how the brain works. The actual structure- how much space is taken up by different areas- is vitally important, as is the density of the neurons. Tyrannosaurus, for instance, had a fairly undeveloped cerebral cortex, suggesting that, although its senses were finely attuned, it was probably not a better planner than other large theropods. From ornithomimids onwards, the brain takes on a conspicuously more bird-like shape, filling the braincase enough to notably change the shape of the skull roof (Holtz). And although bird brains are smaller than those of comparably sized mammals, they pack around twice as many neurons into the same space (Caspar et al).
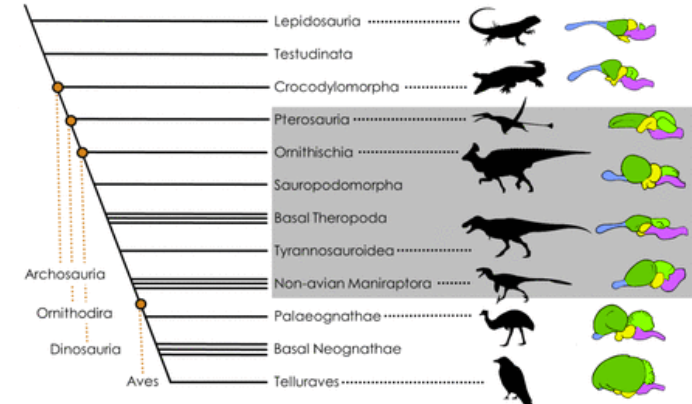
Bird brain evolution (Caspar et al).
One controversial study (Herculano-Huzel) has claimed that, assuming a bird-like neural density, Tyrannosaurus’ brain size alone would make its neuron count comparable to that of a baboon. Other researchers (Caspar et al) are highly skeptical, arguing that its primitive brain structure corresponds to a lower, more croc-like neuron density; that the brain would not have taken up nearly as much of its casing as she estimates; and that the change to a more bird-like count only started with these more advanced forms.
Oddly Familiar Ornithomimosaurs
It's an old cliche that ornithomimosaurs looked just like ostriches, but with arms instead of wings. In fact, thanks to a remarkable series of discoveries from the sandstones of Alberta (previously thought unable to preserve such fine impressions), we now know they even had wings on their arms! Like in ostriches, these long plumes only showed up once they reached adulthood, suggesting they were used for brooding or attracting mates (Reest et al; Zelenitsky). They were probably highly invested in social signaling, because, as the most common small theropods of the end-Cretaceous, they're regularly found in mixed-age groups (Kobayashi and Lu), sometimes of different species (Chinzorig et al). (Because more energy gets used up with each trophic level, herbivores tend to be much more common than carnivores).
Did they eat like ostriches, too? Since their discovery, it's been hypothesized that they fed from branches, using their slender, rakelike hands to hook down branches, and their beaks to nip off leaves (Osborn). These jaws were fairly weak, and they probably did most of the processing in the gut, aided by swallowing stones (Button and Zanno). Like their modern counterparts, they probably ate anything they could get. Some have suggested, based on ridges in the upper jaw, that they could've even filter-fed like ducks, although these were more probably used to crop tough vegetation (Barrett). One weird exception is Early Cretaceous Pelecanimimus. Like other theropods, it retained teeth, but there were over 200 of them! These were tiny and unserrated, forming a continuous surface that, together with its pelican-like pouch, might have been used to trap fish (Perez-Moreno et al).
But the weirdest must be Deinocheirus. For over 50 years, its only known material was an 8-foot pair of arms, suggesting a “bird-mimic” the size of the biggest tyrannosaurs! Finally, in 2013, two specimens, preserving the whole skeleton between them, were reassembled after being broken apart by poachers and shipped to black markets around the world (Holtz). It was not only big, it was highly specialized, with a broad, duck-like bill and a deep, surprisingly under-muscled lower jaw for shoveling soft water plants (one specimen was even found with hundreds of tiny fish bones in its pile of gastroliths!) Supporting its massive size meant Deinocheirus had to give up on the running adaptations of its smaller relatives. Its legs were short and stocky, with broad feet, and its hips were massive, supporting a set of back-stiffening spines almost as tall as Spinosaurus’ (Lee et al.)! Finally, despite retaining its more advanced shape, the brain was tiny, and its sense of balance and coordination were poorly developed.

4-meter Ornithomimus vs 11-meter Deinocheirus (Hartman).
More Mystery Arms
Therizinosaurs are another group with a convoluted history. When the similarly massive arms of Therizinosaurus were described in 1954, they were thought to belong to a giant turtle! Early speculations on the identity of their bits and pieces ranged from enormous predators to insectivorous diggers, fish-eating waders, and, most popularly, highly specialized, late-surviving prosauropods (Barsbold and Perle; Paul). Look at a derived therizinosaur, and you'll see why: the tiny head, perched on a long neck, was heavily modified for cropping and chewing, with densely packed, leaf-shaped teeth and a toothless beak at the front (Button and Zanno; Lautenschlager et al). The whole body was built to accommodate massive guts, including wide hips with a backward-pointing pubis. Finally, to support that weight, they had short, wide feet with that most un-theropod-like of features: four fully functional, weight-bearing toes. It took the discovery of more primitive forms (Zanno), as well as better specimens (Clark et al), to identify them for what they really were: the most committed herbivores among the meat-eating dinosaurs.

Beipiaosaurus (Headden) vs a derived therizinosaur (Paul).
One of these discoveries, the more typically proportioned Beipiaosaurus, was among the first Yixian dinosaurs to preserve evidence of feathers, including an entirely unique set of long, stiff monofilaments (Xu et al). Another set of coelurosaurian features comes from the braincase: like other big-brained theropods, they retained well-developed senses of smell, hearing, and balance, perhaps repurposing them for foraging or social behavior (Lautenschlager 2012).
Therizinosaur means “scythe lizard”, but a common nickname is “sloth dinosaurs” (Holtz). Like Ice-Age ground sloths, they would've used their long, powerful arms and giant claws to strip leaves and pull down branches. In Therizinosaurus, these claws were a full meter long! With the long neck and vertical body, these would've helped extend their feeding range, aided by heavily modified wrists and shoulder-girdles that gave them an exceptional range of motion (Qin et al; Lautenschlager). Of course, they probably served other functions, like display or defense, too.
Another case of weird arms and mistaken identities comes from the much smaller, single-clawed alvarezsaurs. Like therizinosaurs, the most common, specialized end-members were discovered first, and only after did we get a good look at their evolution. When Mononykus (from the same formation as both Deinocheirus and Therizinosaurus!) was first described, features like the backward-pointing pubis; the massive, deeply keeled breastbone; the highly fused, simplified skull and hands; and the hypergracile legs made researchers (Perle et al) believe it was some kind of weird flightless bird. Later discoveries of more primitive forms showed these were convergent adaptations. But for what? They were obviously fast runners, and their weak skulls and tiny teeth suggest they were probably going after much smaller prey. The breastbone would've supported huge muscles, and the claws, though bizarrely tiny, were robust enough to make powerful punctures or support the strains of digging (Qin et al). Finally, their large eyes and long cochleae suggest they were exceptional nocturnal hunters (Choiniere et al). They may have laid flat against a tree trunk or insect colony, listening for motion within, keeping an eye out for predators while poising their arms for a break-in.

Haplocheirus (Choiniere et al) vs Mononykus (Hartman).
Finally, it's important to note that, although these theropods are all typically Cretaceous groups, the presence of more derived bird-like forms in the Jurassic means they must have also been present alongside them. Indeed, we know from direct evidence that there were Jurassic alvarezsaurs. Haplocheirus looks like a typical small theropod, but it already shows the beginning of a robust thumb-claw and a mouth full of tiny teeth (Choiniere 2010). The holotype was 2 meters long and not yet full-grown, suggesting that as alvarezsaurs became specialized for hunting tiny prey, they got smaller themselves (Qin et al).
Like other groups of coelurosaurs, alvarezsaurs probably originated in Asia. But unlike therizinosaurs, which were mainly restricted there (with rare North American exceptions), they seem to have been present wherever there were nests to raid. Evolving such a wide range of dietary specializations meant that coelurosaurs were able to spread and diversify in ways that other theropods had previously been unable to. With new varieties of plant life producing new ecological interactions, the Cretaceous world was becoming more complex, and they were poised to take full advantage of it. But when this complex ecosystem eventually collapsed, this dietary flexibility would prove to have another advantage.
Image Credits
Skeletals by Scott Hartman from skeletaldrawing.com/theropods
Beipiaosaurus by Jaimie Headden from Wikimedia Commons
Paul, G. S. (2016). The Princeton Field Guide to Dinosaurs. Princeton University Press.
Works Cited
Michael J. Benton, Peter Wilf, Hervé Sauquet (2021). The Angiosperm Terrestrial Revolution and the Origins of Modern Biodiversity. New Phytologist, 233(5). doi.org/10.1111/nph.17822
Gurung, K., Field, K. J., Batterman, S. A., Goddéris, Y., Donnadieu, Y., Porada, P., Taylor, L. L., & Mills, B. J. (2022). Climate windows of opportunity for plant expansion during the phanerozoic. Nature Communications, 13(1). https://doi.org/10.1038/s41467-022-32077-7
Lindsay E. Zanno and Peter J. Makovicky (2010). Herbivorous ecomorphology and specialization patterns in theropod dinosaur evolution. Proceedings of the National Academy of Sciences; DOI: 10.1073/pnas.1011924108
Caspar, K. R., Gutiérrez-Ibáñez, C., Bertrand, O. C., Carr, T., Colbourne, J., Erb, A., George, H., Holtz, T. R., Naish, D., Wylie, D. R., & Hurlburt, G. R. (2024). How Smart Was T. Rex? Testing Claims of Exceptional Cognition in Dinosaurs and the Application of Neuron Count Estimates in Palaeontological Research. bioRxiv (preprint). https://doi.org/10.1101/2024.01.10.575006
Herculano-Houzel S. (2023) Theropod dinosaurs had primate-like numbers of telencephalic neurons. Comparative Neurology. 531(9). doi:10.1002/cne.25453.
Hurlburt, Grant; Ridgley, R., Witmer, L. (2013) Relative Size of Brain and Cerebrum in Tyrannosaurid Dinosaurs: An Analysis Using Brain-Endocast Quantitative Relationships in Extant Alligators. In book: Origin, Systematics, and Paleobiology of the Tyrannosauridae. J. M. Parrish, M. Henderson, P. J. Currie, and E. Koppelhus (Eds.)
Van Der Reest, Aaron J.; Wolfe, Alexander P.; Currie, Philip J. (2016). "A densely feathered ornithomimid (Dinosauria: Theropoda) from the Upper Cretaceous Dinosaur Park Formation, Alberta, Canada". Cretaceous Research. 58. doi:10.1016/j.cretres.2015.10.004.
Zelenitsky, D. K.; Therrien, F.; Erickson, G. M.; DeBuhr, C. L.; Kobayashi, Y.; Eberth, D. A.; Hadfield, F. (2012). "Feathered non-avian dinosaurs from North America provide insight into wing origins". Science. 338 (6106). doi:10.1126/science.1225376.
Kobayashi, Y.; Lü, J.-C. (2003). "A new ornithomimid dinosaur with gregarious habits from the Late Cretaceous of China". Acta Palaeontologica Polonica. 48 (2).
Chinzorig, T.; Kobayashi, Y.; Saneyoshi, M.; Tsogtbaatar, K.; Batamkhatan, Z.; Ryuji, T. (2017). "Multitaxic bonebed of two new ornithomimids (Theropoda, Ornithomimosauria) from the Upper Cretaceous Bayanshiree Formnation of southeastern Gobi desert, Mongolia". Journal of Vertebrate Paleontology. Program and Abstracts: 97.
Osborn, Henry Fairfield (1917). "Skeletal adaptations of Ornitholestes, Struthiomimus, Tyrannosaurus". Bulletin of the American Museum of Natural History. 35: 733–771.
Button, D. J.; Zanno, L. E. (2019). "Repeated Evolution of Divergent Modes of Herbivory in Non-avian Dinosaurs". Current Biology. 30(1). doi:10.1016/j.cub.2019.10.050
Barrett, P. M. (2005). "The diet of ostrich dinosaurs (Theropoda: Ornihomimosauria)". Palaeontology. 48 (2). doi:10.1111/j.1475-4983.2005.00448.x
Pérez-Moreno, B. P.; Sanz, J. L.; Buscalioni, A. D.; Moratalla, J. J.; Ortega, F.; Rasskin-Gutman, D. (1994). "A unique multitoothed ornithomimosaur dinosaur from the Lower Cretaceous of Spain". Nature. 370 (6488). doi:10.1038/370363a0.
Holtz, Thomas (2014). "Paleontology: Mystery of the horrible hands solved". Nature. 515 (7526). doi:10.1038/nature13930.
Lee, Y.N.; Barsbold, R.; Currie, P.J.; Kobayashi, Y.; Lee, H.J.; Godefroit, P.; Escuillié, F.O.; Chinzorig, T. (2014). "Resolving the long-standing enigmas of a giant ornithomimosaur Deinocheirus mirificus". Nature. 515 (7526). doi:10.1038/nature13874.
Barsbold, R.; Perle, A. (1980). "Segnosauria, a new infraorder of carnivorous dinosaurs". Acta Palaeontologica Polonica. 25 (2).
Paul, G. S. (1984). "The segnosaurian dinosaurs: relics of the prosauropod-ornithischian transition?". Journal of Vertebrate Paleontology. 4 (4). doi:10.1080/02724634.1984.10012026
Lautenschlager, S.; Lawrence, M. W.; Perle, A.; Zanno, L. E.; Emily, J. R. (2014). "Cranial anatomy of Erlikosaurus andrewsi (Dinosauria, Therizinosauria): new insights based on digital reconstruction". Journal of Vertebrate Paleontology. 34 (6). doi:10.1080/02724634.2014.874529.
Zanno, L. E. (2010). "Osteology of Falcarius utahensis (Dinosauria: Theropoda): characterizing the anatomy of basal therizinosaurs". Zoological Journal of the Linnean Society. 158 (1). doi:10.1111/j.1096-3642.2009.00464.x.
Clark, J. M.; Perle, A.; Norell, M. (1994). "The skull of Erlicosaurus andrewsi, a Late Cretaceous ‘Segnosaur’ (Theropoda, Therizinosauridae) from Mongolia'. American Museum Novitates (3115)
Qin, Z., Liao, CC., Benton, M.J. et al. (2023). Functional space analyses reveal the function and evolution of the most bizarre theropod manual unguals. Commun Biol 6(181). https://doi.org/10.1038/s42003-023-04552-4
Lautenschlager, S. (2014). "Morphological and functional diversity in therizinosaur claws and the implications for theropod claw evolution". Proceedings of the Royal Society B. 28 (1785). doi:10.1098/rspb.2014.0497.
Xu, X.; Zheng, X.; You, H. (2009). "A new feather type in a nonavian theropod and the early evolution of feathers". Proceedings of the National Academy of Sciences. 106 (3). doi:10.1073/pnas.0810055106
Lautenschlager, S.; Rayfield, E. J.; Perle, A.; Zanno, L. E.; Witmer, L. M. (2012). "The Endocranial Anatomy of Therizinosauria and Its Implications for Sensory and Cognitive Function". PLOS ONE. 7 (12): e52289. doi:10.1371/journal.pone.0052289
Perle, A.; Chiappe, L. M.; Rinchen, B.; Clark, J. M.; Norell, M. A. (1994). "Skeletal morphology of Mononykus olecranus (Theropoda, Avialae) from the late Cretaceous of Mongolia". American Museum Novitates (3105)
Choiniere, J.N.; Neenan, J.M.; Schmitz, L.; Ford, D.P.; Chapelle, K.E.J.; Balanoff, A.M.; Sipla, J.S.; Georgi, J.A.; Walsh, S.A.; Norell, M.A.; Xu, X.; Clark, J.M.; Benson, R.B.J. (2021). "Evolution of vision and hearing modalities in theropod dinosaurs". Science. 372 (6542). doi:10.1126/science.abe7941
Choiniere, J. N.; Xu, X.; Clark, J. M.; Forster, C. A.; Guo, Y.; Han, F. (2010). "A basal alvarezsauroid theropod from the Early Late Jurassic of Xinjiang, China". Science. 327 (5965): 571–574. Bibcode:2010Sci...327..571C. doi:10.1126/science.1182143
Qin, Zichuan; Zhao, Qi; Choiniere, Jonah N.; Clark, James M.; Benton, Michael J.; Xu, Xing (2021-07-06). "Growth and miniaturization among alvarezsauroid dinosaurs". Current Biology. 31 (16).e5. doi:10.1016/j.cub.2021.06.013




